
If you’ve never heard of Spathaspora passalidarum, you’re not alone. This tongue-twisting yeast has been quietly loitering in the shadow of industrial giants like Saccharomyces cerevisiae (S. cerevisiae)—until now. A new study has shined the lab light on this low-key microbe, and what it reveals might just shake up how we think about second-generation biofuels.
Let’s back up for a second. Second-gen bioethanol—the kind made from agricultural leftovers like corn stalks, wheat straw, or sugarcane bagasse—is the sustainability dream that keeps running into metabolic reality. These byproducts are loaded with a messy mix of sugars, and our usual microbial workhorses just aren’t up for the challenge. But S. passalidarum? It might be.
This isn’t the kind of yeast that parties with just glucose and calls it a night. It’s got a more diverse palate—and a few quirks that bioengineers, fermentation nerds, and climate optimists should all be paying attention to.

🧪 The Sugar Buffet Test: Mixed Plates, Mixed Results
In this study, S. passalidarum was invited to a metabolic all-you-can-eat buffet: hexoses, pentoses, disaccharides, sugar alcohols. Think: glucose, galactose, xylose, fructose, mannose, maltose, arabinose, glycerol—even lactose, which turned out to be the yeast equivalent of a snubbed salad at a steakhouse.
The results weren’t exactly uniform. Hexoses like glucose, galactose, and mannose were devoured with gusto, yielding a solid return of ethanol. Maltose joined the high-performance club too. Meanwhile, arabinose and glycerol were metabolized—just not for ethanol. They went toward biomass, or in arabinose’s case, arabitol. That’s a sugar alcohol, which isn’t useless, but it’s not what you pump into a bioethanol tank.
And lactose? Completely ignored. Sorry, dairy.
🔄 Sugar Politics: It’s Not Just What You Eat, But When
Now here’s where things get interesting—and a little neurotic. When S. passalidarum is working on xylose (a major sugar in plant biomass) and suddenly hexoses like glucose or galactose show up, it just… stops. Like someone mid-conversation dropping everything when cake enters the room. This isn’t just preference—it’s inhibition. Even non-metabolizable sugars like 2-deoxyglucose triggered the pause, suggesting early-stage regulation at the sugar uptake or phosphorylation level.
We call this the “sugar hierarchy problem.” And it’s not trivial. If you’re designing an industrial process to ferment biomass, sugar order and feeding strategy can make or break your yield timeline.
Strangely, fructose and sucrose didn’t cause this delay. Which means… there’s a loophole. And if there’s a loophole, there’s a process strategy waiting to be born.

🌬️ Oxygen: Not Just for Humans
The sugar story flips again when oxygen enters the chat.
In aerobic conditions, arabinose—useless under fermentation—gets converted into arabitol and biomass. Glycerol use also improves. Still no ethanol, but it hints at value in bioproduct diversification. So while these substrates might not be good fuel makers, they could be side hustles: biomass for protein, arabitol for food or pharma.
Translation: ethanol may not be the only ROI worth chasing.
🚧 The Industrial Red Flags
Before we get too excited, let’s get real. S. passalidarum is no magic wand.
It doesn’t tolerate ethanol well. In a production tank, that’s like a chef who gets dizzy in a kitchen.
It’s sensitive to inhibitors—furfural, HMF, and acetate—all of which are common in biomass hydrolysates.
The study didn’t even touch on gene expression, sugar transporters, or metabolic engineering. That’s a whole unexplored continent.
But none of these are deal-breakers. They’re checklists. And for microbial engineers, checklists are invitations.
🔬 What the Study Means for Biotech
So what’s the practical takeaway here?
Strain Engineering Is Inevitable
Want better ethanol tolerance or detox pathways? Time to get into the genome.
Process Design Must Get Smarter
Feed sugars in sequence, run detox steps, or build microbial teams. Co-cultures could help balance sugar types and byproduct tolerances.
Valorize Side Products
Don’t just chase ethanol. Arabitol or biomass have real market value in food tech, materials, and fine chemicals.
Learn the Regulation Game
Cracking the sugar uptake hierarchy could lead to smarter fermentation controls—and better yields, faster.
Test with Real Biomass
The study used pure sugars. That’s nice, but the real world is messier. Let’s see how S. passalidarum performs in actual bagasse soup.
🍷 So, Is Spathaspora Worth the Hype?
She’s not flawless. She’s not fast. She’s not even that famous. But Spathaspora passalidarum is a rare yeast that offers metabolic range, sugar flexibility, and a quiet kind of resilience. She might not replace S. cerevisiae on the fermentation throne anytime soon—but she doesn’t have to. What she offers is niche power.
In the evolving drama of climate change and renewable energy, we don’t just need one hero microbe. We need an ensemble cast. S. passalidarum may be the indie darling, the supporting actor with surprising depth, the one who—once you give her the right stage—could deliver a performance that shifts the storyline.
Because in the biofuel world, turning waste into power isn’t just about fermentation. It’s about flexibility. And in that department, this shadow-dwelling yeast just might shine.
Key Takeaways
- Spathaspora passalidarum is a native North American yeast discovered in the guts of wood-boring beetles that possesses an unusual ability to ferment xylose—the primary sugar in agricultural lignocellulosic waste—into ethanol, a capability rare among yeasts.
- Xylose fermentation is the key bottleneck in converting agricultural residues (corn stover, wheat straw, sugarcane bagasse) to fuel ethanol: these feedstocks contain 20–35% xylose that industrial yeast strains (Saccharomyces cerevisiae) cannot efficiently ferment.
- S. passalidarum’s natural xylose fermentation pathway involves a single xylose reductase with cofactor flexibility, avoiding the redox imbalance problems that make engineered xylose fermentation in S. cerevisiae inefficient.
- Identifying and engineering superior xylose-fermenting yeasts is critical to the economics of second-generation (cellulosic) biofuels, which could use agricultural waste rather than food-competing feedstocks like corn grain.
- The discovery of S. passalidarum exemplifies bioprospecting—the search for naturally evolved organisms with industrially valuable metabolic capabilities—as an approach to solving biochemical engineering challenges that synthetic biology alone has not yet overcome.
Frequently Asked Questions
What makes Spathaspora passalidarum special as a yeast for biofuels?
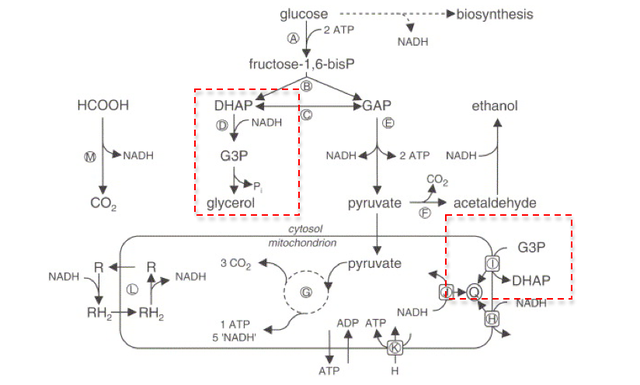
Spathaspora passalidarum is remarkable among yeasts for its native ability to efficiently ferment xylose—the second most abundant sugar in plant cell walls—into ethanol without genetic engineering, overcoming the primary metabolic limitation of industrial yeast strains. The industrial significance of xylose fermentation: plant cell walls (lignocellulosic biomass) contain two major sugar components: cellulose (polymer of glucose) and hemicellulose (complex polymer containing primarily xylose, arabinose, and other pentose sugars). In agricultural residues, xylose typically represents 20–35% of total fermentable sugars; in corn stover: approximately 22% xylose; in wheat straw: approximately 20–28% xylose; in sugarcane bagasse: approximately 21–26% xylose. This means that a biofuel process that cannot ferment xylose leaves 20–35% of the feedstock’s sugar value unused. Standard industrial yeast (Saccharomyces cerevisiae): does not naturally ferment xylose at industrially useful rates; intensive metabolic engineering over decades has produced xylose-fermenting S. cerevisiae strains; however, engineered S. cerevisiae still ferments xylose far less efficiently than glucose—slower rates, lower yields, and sensitivity to inhibitors. What makes S. passalidarum exceptional: discovered in the hindgut of Odontotaenius disjunctus beetles living in decaying wood; in its natural habitat, it evolved to ferment xylose (abundant in wood hemicellulose) efficiently; naturally produces xylose reductase (XR) and xylitol dehydrogenase (XDH) enzymes that are unusually well-balanced in their cofactor preferences, avoiding the xylitol accumulation problem that limits engineered S. cerevisiae.
Why can’t regular yeast ferment the sugar xylose?
The inability of Saccharomyces cerevisiae—the workhorse of industrial fermentation globally—to efficiently ferment xylose reflects the evolutionary trajectory of a yeast specialised for glucose-rich fruit environments that never needed xylose catabolism, combined with a specific biochemical problem that is difficult to solve through simple genetic engineering. Why S. cerevisiae doesn’t ferment xylose: evolutionary context: S. cerevisiae evolved in sugar-rich environments (rotting fruit, flower nectar) where glucose and fructose are the primary sugars; it never encountered lignocellulosic biomass as a primary food source; it therefore never evolved the enzymatic machinery for efficient pentose sugar metabolism. The biochemical barrier—cofactor imbalance: xylose fermentation to ethanol requires a two-step oxidation-reduction sequence in the fungal pathway: xylose → xylitol (catalysed by xylose reductase/XR) → xylulose (catalysed by xylitol dehydrogenase/XDH) → ethanol (via phosphoketolase pathway). The problem: XR in most fungi uses NADPH as its preferred electron donor; XDH uses NAD⁺ as its electron acceptor; under anaerobic conditions, the cell has excess NADH and limited NAD⁺; XDH cannot oxidise xylitol to xylulose efficiently when NAD⁺ is limiting; xylitol accumulates instead of being converted to xylulose and then ethanol; in aerobic conditions, the cofactor balance is maintained, but aerobic fermentation is inefficient for ethanol production (ethanol is oxidised to CO₂ in excess oxygen). S. passalidarum’s solution: its XR enzyme has dual cofactor specificity (uses both NADH and NADPH), unlike most fungi where XR strongly prefers NADPH; this cofactor flexibility maintains redox balance under the anaerobic conditions needed for efficient ethanol production.
How close are we to commercial cellulosic ethanol and what are the remaining challenges?
Commercial cellulosic ethanol has been in development for decades and several commercial-scale facilities have been built, but persistent technical and economic barriers have prevented the technology from becoming cost-competitive with first-generation grain ethanol or fossil fuels without continued subsidy support. Current status: commercial plants: several large commercial cellulosic ethanol plants were built in the US between 2012 and 2015—POET-DSM Project Liberty (Iowa), Abengoa Bioenergy (Kansas), DuPont Cellulosic Ethanol (Iowa)—using corn stover as feedstock; almost all have encountered severe economic challenges: DuPont’s plant was sold and shut down; Abengoa’s plant closed after the parent company’s bankruptcy; POET-DSM’s plant operated at reduced capacity; technical challenges in scale-up exceeded projections and production costs remained substantially above market ethanol prices. Economic barriers: feedstock costs and logistics—collecting, storing, and transporting bulky, low-density agricultural residues (corn stover, wheat straw) to a central processing facility is expensive; logistics favour smaller distributed facilities but chemical conversion achieves economies of scale at larger plant sizes. Pretreatment cost—breaking down crystalline cellulose and removing hemicellulose to make sugars accessible requires high-temperature acid or steam explosion pretreatment followed by enzyme addition; enzyme costs (cellulases) have fallen substantially but remain significant. Inhibitor toxicity—pretreatment generates fermentation inhibitors (furfural, hydroxymethylfurfural, acetic acid) that stress fermenting organisms; robust tolerant strains are needed. Xylose fermentation—where S. passalidarum research is relevant; current commercial processes either waste xylose or ferment it inefficiently. Current technological readiness: lignocellulosic conversion is technically feasible; the question is economic competitiveness; current production costs estimated at USD 0.90–1.50 per litre, versus approximately USD 0.40–0.60 for first-generation corn ethanol; new strain development incorporating superior xylose fermenters like S. passalidarum characters could meaningfully reduce cost gaps.
What other organisms can ferment xylose and how do they compare to S. passalidarum?
The ability to ferment xylose efficiently has evolved independently in multiple lineages of fungi and bacteria, and comparative evaluation of these diverse organisms has been a productive area of industrial biotechnology research. Natural xylose-fermenting organisms: yeasts: Scheffersomyces stipitis (formerly Pichia stipitis)—the most extensively studied and used native xylose-fermenting yeast prior to S. passalidarum; isolated from wood-inhabiting insects; excellent xylose fermenter but requires specific low-oxygen conditions and is sensitive to ethanol; XR/XDH pathway with partially dual cofactor specificity. Candida shehatae—another naturally xylose-fermenting yeast found in wood-boring insect guts; similar to S. stipitis in properties; less studied for industrial applications. Spathaspora species—a genus of yeasts predominantly found in beetle gut environments in decaying wood; S. passalidarum emerged from screening within this genus as having superior characteristics; other Spathaspora species are also being evaluated. Bacteria: Zymomonas mobilis—naturally ferments glucose to ethanol very efficiently (via Entner-Doudoroff pathway); has been engineered by NREL to incorporate xylose fermentation genes (xylA, xylB, tal, tkt); engineered Z. mobilis can ferment xylose but typically requires further optimisation for mixed substrate fermentation. Escherichia coli—engineered strains can ferment xylose (and other pentose and hexose sugars) through metabolic engineering; less tolerance to ethanol than yeast limits viability. Comparative advantages of S. passalidarum: high native xylose fermentation rates under oxygen-limited conditions; produces ethanol rather than xylitol from xylose; better cofactor balance than S. stipitis in anaerobic conditions; some ethanol tolerance; being evaluated as a stand-alone fermenter and as a gene donor for S. cerevisiae engineering. Limitations of S. passalidarum: less ethanol tolerant than industrial S. cerevisiae (ethanol tolerance approximately 40 g/L vs. 100+ g/L for robust S. cerevisiae strains); less inhibitor tolerant than industrial S. cerevisiae strains; not yet as genetically tractable (tools for genetic manipulation less developed than for S. cerevisiae).
Could yeast-based biofuels from agricultural waste really replace significant amounts of fossil fuel?
The potential of lignocellulosic yeast fermentation to produce ethanol from agricultural residues at scale sufficient to meaningfully displace fossil fuel use is a question of resource availability, land use, technical feasibility, and energy economics—with both real potential and important caveats. Resource availability estimate: global agricultural residue production: approximately 3.8–4.5 billion dry tonnes of agricultural residues are generated annually (FAO estimates); a fraction of this is practically collectible without damaging soil health (roughly 30–50% depending on region); collectible lignocellulosic residue might represent 1.1–2.0 billion dry tonnes globally. Theoretical ethanol yield: approximately 180–280 litres of ethanol per dry tonne of lignocellulosic biomass in optimised processes (lower values with current processes); at 200 litres per dry tonne from 1.5 billion dry tonnes: 300 billion litres of ethanol; global gasoline consumption is approximately 1,800 billion litres equivalent annually. Theoretical replacement: agricultural residues alone could theoretically supply approximately 15–20% of global gasoline equivalent if all were converted optimally—substantial but not a full replacement. Real-world limitations: soil carbon maintenance: removing too much crop residue depletes soil organic matter, increasing erosion and reducing soil productivity; sustainable residue harvest limits mean practically less than 30% of residues in most cropping systems. Net energy balance: the energy return on energy investment (EROI) for cellulosic ethanol is higher than for corn grain ethanol (a controversial point) but requires validation at commercial scale. Infrastructure: liquid fuel distribution infrastructure favours ethanol blending (E10, E15) or high-blend markets (Brazil E100 motorcycles); transitions to higher blends require vehicle compatibility. The role of superior xylose fermenters: better xylose fermentation (where S. passalidarum-derived improvements contribute) can improve ethanol yield per tonne of feedstock by 15–30% by capturing previously wasted xylose—potentially significant for the economics and efficiency of the entire sector.







